Peptides can engage the large, shallow biomolecular surfaces that small molecules struggle to reach, yet rapid proteolytic degradation in biological environments limits their use as probes and therapeutics. Endogenous proteases process canonical α-peptide backbones because those backbones tend to adopt β-strand-like conformations before cleavage. Chemists have developed macrocyclization, noncanonical residue incorporation, and backbone amide substitution to counter this vulnerability. Backbone N-methylation remains an extensively studied approach, with documented positional effects on proteolytic susceptibility, yet N-methylation can impose conformational penalties that undermine peptides whose activity depends on extended backbone geometry. The field needs backbone editing tools with a lighter conformational footprint.
Researchers in the Del Valle Group at the University of Notre Dame, published in RSC Chemical Biology, evaluated backbone N-amino and N-hydroxy substitution as an alternative. Working from a defined chymotrypsin substrate, the team replaced individual residues from positions P3 through P3′ with their corresponding α-hydrazino acid counterparts, using an oxaziridine amination reagent and standard Fmoc SPPS. They then prepared N-hydroxy peptides through a hydroxamate-mediated ligation and N-methyl peptides by conventional SPPS. Chymotrypsin degradation assays monitored by analytical RP-HPLC yielded half-lives from single-phase exponential decay fits.
Position governed the outcome. An α-hydrazino acid at P1, immediately before the scissile bond, or at P1′, the first residue after it, extended half-lives 100- to 150-fold over the unmodified parent, while modifications farther from the cleavage site stayed within one order of magnitude of the control. At P1, N-hydroxylation raised stability roughly 40-fold, below the corresponding N-amino and N-methyl variants but still a clear gain. At P1′, hydroxamate substitution offered only modest protection, likely because the electrophilic hydroxamate carbonyl at the scissile bond hydrolyzes more readily than its hydrazide and tertiary amide counterparts. Circular dichroism spectra showed random coil signatures across the series, indicating that these stability differences do not arise from global conformational changes, though CD cannot resolve local effects at the scissile bond.
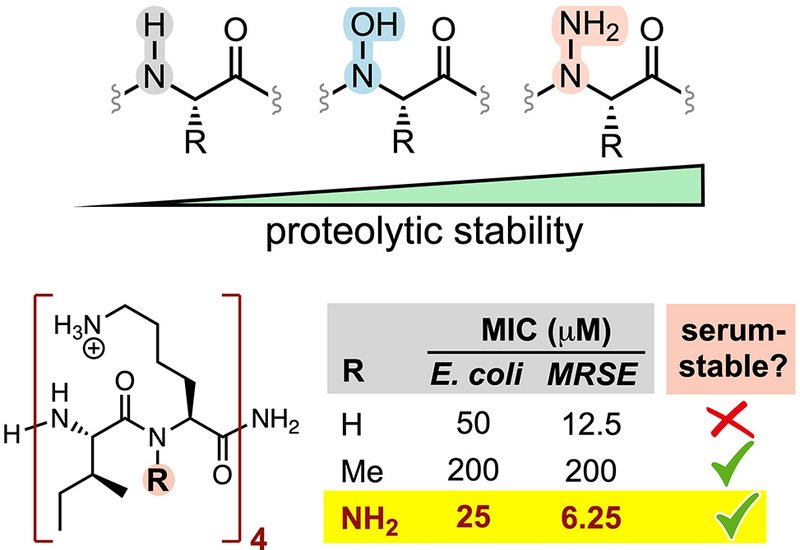
The team then asked whether N-amination could stabilize a conformationally active peptide without destroying its function. They chose a β-sheet-forming antimicrobial octapeptide, H-[Ile-Lys]4-NH2, whose amphipathic structure disrupts membranes in Escherichia coli and methicillin-resistant Staphylococcus epidermidis. Replacing Lys residues with α-hydrazino Lys maintained comparable antibacterial activity, and the tetra-N-aminated analogue, peptide 16, doubled activity against both pathogens. N-methylation of two or more residues reduced membrane lytic activity, consistent with its known conformational penalties in amphipathic β-sheet peptides. Peptide 16 remained intact after 24 hours of trypsin exposure and after 400 hours in 25% human serum, conditions under which the parent degraded within 20 minutes and 3 hours. CD spectroscopy in SDS micelles showed that tetra-N-amination preserved environment-dependent folding, with cooperative thermal denaturation near 60 °C marking a structured membrane-associated state despite an unnatural residue content of 50%.
These results position backbone N-heteroatom substitution as a chemically distinct strategy that separates proteolytic protection from the conformational disruption of N-methylation. The approach suits peptides whose function depends on extended or β-sheet-like geometry, a class poorly served by existing methods. The Del Valle group expects N-amination and N-hydroxylation to aid the design of folded peptide and protein mimics for cellular and in vivo applications, widening the toolkit for protease-resistant probes and candidate therapeutics.