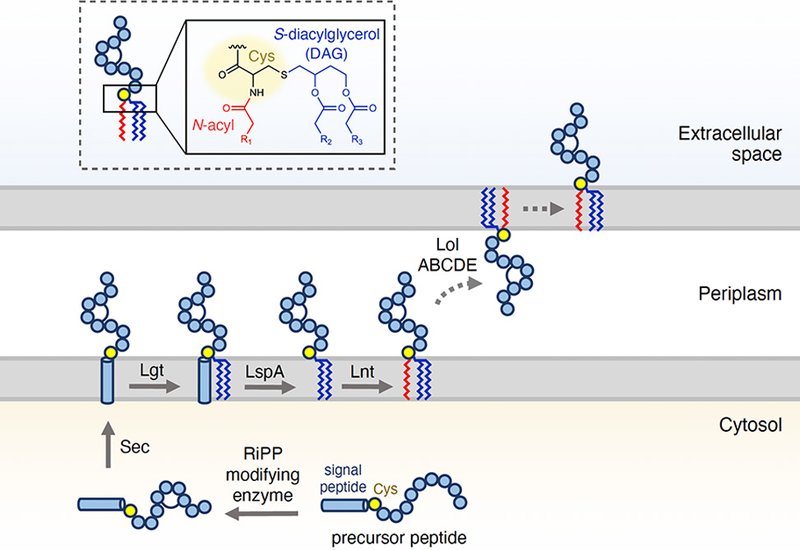
Ribosomally synthesized and posttranslationally modified peptides, RiPPs, encompass antibiotics, antifungals, and metal-binding metallophores whose chemical diversity arises from the action of dedicated modifying enzymes on a precursor peptide. One recently characterized enzyme, ChrH, belongs to the multinuclear non-heme iron-dependent oxidative enzyme, MNIO, family and installs a macrocycle, a hydantoin heterocycle, and an S-methyl group onto its substrate ChrA. Whether closely related MNIOs produce structurally distinct products, and whether any of those products undergo maturation outside the cytoplasm, remained open questions. The bacterial lipoprotein biosynthesis pathway is ubiquitous in gram-negative bacteria and uses dedicated enzymes to attach diacylglycerol, DAG, and acyl groups to an N-terminal cysteine residue, but no RiPP had previously been shown to exploit this route.
Researchers in the van der Donk Group at the University of Illinois at Urbana-Champaign, published in PNAS, used comparative genomics to identify ChrH homologs across Bacteroidota, Cyanobacteria, and Myxococcota and selected representative members for biochemical characterization. Heterologous coexpression of precursor peptides with their cognate MNIOs in Escherichia coli, followed by MALDI-TOF MS, NMR spectroscopy, and tandem MS, established the chemical structures of the modified products. SignalP 6.0 analysis revealed that several precursors carry N-terminal lipoprotein signal peptides, prompting isolation and characterization of the lipidated forms by immobilized metal affinity chromatography and on-membrane LysC digestion.
Coexpression of DybA from Dyadobacter crusticola with the MNIO DybH produced a macrocyclic, heterocycle-containing product bearing a free, unmethylated thiol, a −4 Da shift relative to the unmodified precursor that NMR spectroscopy confirmed as a thiohemiaminal macrocycle lacking the S-methyl group present in ChrAH. Comparative experiments across Bacteroidota and Cyanobacteria homologs showed that the ChrH/DybH subfamily tolerates substrates ranging from short peptides to proteins exceeding 34 kDa, including a homodimeric precursor, MelA1, whose C-terminal CPACGMG motif is modified by a two-protein MNIO complex. The broad substrate scope is consistent with relaxed selection pressure on the N-terminal leader region when modification is directed by a minimal C-terminal motif.
Bioinformatic and biochemical evidence demonstrated that DybA contains a canonical SPII lipoprotein signal peptide. After cytosolic modification by DybH, DybA is processed by the endogenous bacterial lipoprotein enzymes: prolipoprotein diacylglyceryl transferase adds a DAG group to the conserved N-terminal cysteine sulfur, signal peptidase II cleaves the signal peptide, and apolipoprotein N-acyltransferase acylates the free amine. MALDI-TOF MS and MS/MS of the purified lipidated product confirmed triacylation, with the predominant species carrying C16:1, C16:0, and C16:0 acyl chains. Genome mining across other MNIO subfamilies, including those responsible for bufferins and oxazolin-containing peptides, revealed additional biosynthetic gene clusters encoding predicted RiPP-lipoprotein hybrids, which the authors designate DAG-RiPPs to distinguish them from N-acylated lipidated RiPPs. A subset of autoinducing peptide precursors was also found to carry predicted SPII signal peptides, extending the potential scope of the pathway beyond MNIOs.
The discovery of DAG-RiPPs establishes a maturation paradigm in which a RiPP precursor exits the cytoplasm for further modification by the membrane-localized lipoprotein machinery, expanding the chemical space accessible to ribosomal biosynthesis. The lipidated products may localize to the inner or outer membrane, or be released in outer membrane vesicles, pointing to potential roles in cell-surface signaling, nutrient acquisition, or interbacterial competition. The authors also note that the macrocyclic core of these products shares structural features with the B-ring of lipid II-binding lantibiotics such as nisin, raising the possibility of antibiotic activity worth investigating. From a biotechnology perspective, RiPP-modifying enzymes could offer a route to posttranslationally diversifying immunogenic lipoprotein adjuvants beyond the canonical 20 amino acids, with implications for vaccine design.