For decades, antimicrobial peptides, AMPs, have stood at the crossroads between innate immunity and drug discovery, yet their molecular diversity remains incompletely explored. Dickey, Otto, and collaborators at the National Institutes of Health, now report in Nature Communications an entirely new AMP family, termed the transmembrane helix–containing bacteriocins, TMcins, that defies long-held assumptions about peptide pore formation. In contrast to classical AMPs that transiently disturb membranes or large protein toxins that assemble stable channels, TMcins achieve both feats within a single short peptide: they oligomerize into discrete, nanometer-scale β-barrel pores stabilized by transmembrane helices.
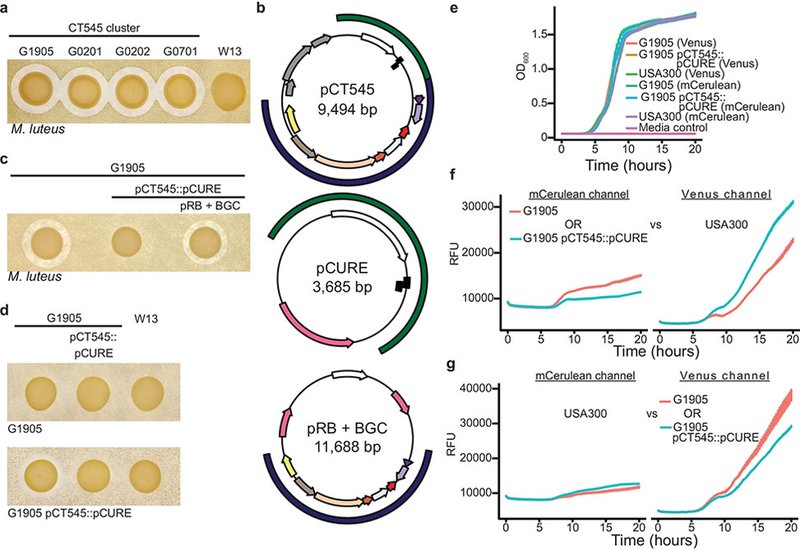
The discovery originated from an unusual cluster of Staphylococcus aureus isolates, CT545, recovered from Buruli ulcer wounds that displayed potent diffusible antimicrobial activity absent in closely related strains. Genomic sleuthing revealed a small plasmid, pCT545, carrying a previously unrecognized biosynthetic gene cluster, BGC whose deletion abolished activity. Complementation restored killing, proving that pCT545 encodes a new ribosomally synthesized AMP. Purification and sequencing identified a 5.8 kDa peptide, TMcin, processed from a 297-bp precursor and secreted via an associated ABC transporter system. Bioinformatic analyses showed that TMcin escaped all current AMP-prediction pipelines, highlighting the limitations of purely computational discovery.
Structural modeling by AlphaFold2 predicted a remarkable architecture: an N-terminal transmembrane α-helix capped by a pair of conserved tryptophans and tethered to a C-terminal two-strand β-sheet loop closed by disulfide bonds. The fold exposes a hydrophilic β-sheet face alongside a hydrophobic helical belt, endowing the peptide with an overall amphipathic character. TMcin homologs carrying this distinctive motif were subsequently identified across multiple Gram-positive genera, including Bacillus and Actinomyces, indicating a broad evolutionary distribution of the family.
Functionally, TMcin-G1905 exhibited potent bactericidal activity against Gram-positive pathogens—methicillin-resistant S. aureus, vancomycin-resistant Enterococcus faecalis, and others—while sparing Gram-negative species, whose outer membranes likely block access. Microscopy and biochemical assays revealed that TMcin associates with cellular membranes, triggers ATP leakage, and dissipates membrane potential without lysing cells outright. The resulting shrunken, depolarized phenotype is consistent with sustained pore formation rather than wholesale membrane dissolution.
Electrophysiology confirmed this mechanistic leap. In planar lipid bilayers, purified TMcin-G1905 produced stepwise current jumps with median conductance of 10.8 nS in 150 mM KCl—orders of magnitude greater than canonical AMP channels such as gramicidin A. The pores displayed linear current–voltage relationships and negligible ion selectivity, signifying large aqueous conduits. AlphaFold2 multimer modeling and microsecond molecular-dynamics simulations converged on stable ring-like assemblies of roughly twenty peptides forming a β-barrel inner wall encircled by a hydrophobic helix belt. Hydrogen bonding and inter-strand salt bridges knit the oligomer together, while simulations in cardiolipin-containing bilayers demonstrated long-lived structural integrity. Circular-dichroism spectra supported mixed α/β content in solution, matching computational predictions.
Functionally, these pores are massive: S. aureus cells exposed to TMcin became permeable to 40 kDa fluorescent dextran and polysucrose probes, confirming translocation channels up to ~10 nm in diameter, comparable to classical protein toxins yet built from a 5.8 kDa peptide. The study further uncovered a natural partnership between TMcin and the staphylococcal surfactant peptides, phenol-soluble modulins, PSMs. PSMs enhanced TMcin solubility and diffusion, acting as molecular chaperones that release the hydrophobic AMP from producer membranes, an elegant co-evolutionary adaptation within the same bacterial species.
Beyond its microbiological novelty, TMcin reshapes our understanding of AMP structure–function space. It bridges the conceptual divide between disordered, transiently permeabilizing peptides and structured, protein-scale pore-formers, establishing a continuum of membrane-active architectures. The TMcin BGC represents a new ribosomally synthesized and post-translationally modified peptide, RiPP, family, previously hidden from genome-mining algorithms due to its unusual sequence and domain composition. Its discovery suggests that additional, unrecognized AMP classes remain embedded within microbial genomes, awaiting phenotype-driven approaches for identification.
By coupling genomics, structural modeling, electrophysiology, and microscopy, Dickey and colleagues reveal an AMP that functions as a true nanoscale pore-forming complex. The implications are profound: TMcins provide a new mechanistic template for engineering stable β-barrel pores from short peptides, opening avenues for synthetic antimicrobial design, biosensing, and controlled molecular transport. At a broader level, they underscore the evolutionary ingenuity of bacterial defense systems and expand the molecular repertoire available to counter antimicrobial resistance.